The startle response of sheep (Krausman and Hervert 1983, Krausman et al. 1998) to high speed military overflights could affect the daily movements, subsequent home range size, and habitat use by sheep. Daily movement might increase if sheep attempted to escape from aircraft disturbance, or movement might decrease if sheep sought out and remained in areas that offered security from disturbance. Home range size might be expected to respond in a similar manner. If daily movements and/or home range area changed substantially as a result of overflights, then changes in habitat use might be expected as well.
Estimating the magnitude and statistical and biological significance of any potential overflight effect is difficult when there is substantial variability in 1) habitats between study areas, 2) annual conditions among years, 3) seasonal conditions within years, and 4) individual sheep. To be deemed statistically significant, sheep response to aircraft activity must be distinguished from natural variance due to habitat, annual and seasonal conditions, and individuals. To be deemed biologically significant, the magnitude of any statistically detectable and/or demonstrated effect must be sufficient to reasonably be expected to affect population performance (e.g. growth, productivity, survival, population size). In both the statistical and biological sense, adequate replication of experimental units (sheep) and treatment levels (i.e. aircraft activity) among classification variables (i.e. study area, year, season within year) is essential for reliable and robust results.
To assess the potential influence of military overflights on sheep behavior and habitat use, we conducted multiple analyses of covariance to estimate whether or not the average number of military flight sorties launched in 2-week periods influenced our estimates of the effect of study area, year within study area, and 2-week period within years within study areas on daily distance moved, home range size, and habitat use of adult female Dall’s sheep in two study areas in Military Operation Areas (MOA) 1 and 2, interior Alaska, from late-April through early-August, 1999-2002.
In two study areas (Cirque Lakes (CL) and West Point (WP)) during four years (1999-2002) approximately 10 adult female sheep were GPS collared in late winter (~March) and their locations recorded at 3 (1999) or 4 (2000-2002) hour intervals for the ensuing 8 months each year. Collars were removed at the end of the monitoring period and a new sample of sheep was instrumented the following year. Analyses were focused on eight successive 2-week periods that began on ~15 April and continued through ~ 10 August each year. These intervals were mutually exclusive within years, and were keyed to actual/scheduled MFEs in each year. As a result, the beginning/ending dates of the 2-week periods were not constant among years, but starting and ending dates for each successive period varied only by ~3 days among years. The periods were named late-April, early-May, Late-May, early-June, late-June, early-July, late-July, and early-August and referred to as sequences 3-10, respectively.
Habitat attributes at locations of sheep (landcover class, slope, aspect, elevation, terrain ruggedness, relative green biomass at peak vegetation development (NDVI; Tucker 1979, Tucker et al. 1986) were derived from remotely sensed TM land cover classifications (Ducks Unlimited 1998) and from digital elevation models. The 18 original TM landcover classes were condensed into forest, shrub-grass-herb, and sparsely vegetated types to reduce the occurrence of empty cells.
NDVI indexes the disproportionate reflectance of near-infrared radiation from green vegetation (Tucker and Sellars 1986) in the canopy of plant communities. We obtained NDVI from a TM image taken on 11 July 1994, the only image which we could find that gave near complete cloud-free coverage of both study areas. NDVI was calculated from bands 3 and 4 as NDVI = (Band4 - Band 3)/(Band4 + Band3) and converted to the decimal range 0-1.0 by setting values less than 0 to 0 (Griffith et al 2002). A portion of the western side of the West Point study area was not covered by the NDVI grid and NDVI was set to missing for that area.
Terrain ruggedness (Nicholson et al. 1997) was estimated as the sum of diversity in slope and diversity in aspect within the average 4-hour-movement radius of locations. We used the sum, rather than the product (Nicholson et al. 1997), of diversity in slope and aspect because the product increased the range of estimated ruggedness approximately 10-fold compared to estimates of ruggedness obtained for slope or aspect alone. The sum maintained the proportional range of the original data.
Movement distance was estimated as the sum of straight line distances between successive locations within 2-week sequential period. This constituted a minimum estimate of total distance traveled.
Minimum convex polygon (MCP) home ranges were calculated from locations for each sheep within 2-week periods. A maximum of 90-120 locations was available for each sheep in each period; MCP home range sizes became asymptotic at approximately 50 locations; sheep with less than 50 relocations for the 2-week sequential periods were excluded from analyses. Usually these were sheep that died or sheep with collar failure within a period.
MCP home ranges were used, in contrast to more sophisticated techniques such as least squares cross validated fixed kernels (Seaman et al. 1996, 1998, 1999), to obtain the simplest reasonable estimate of habitat availability and the extent of area used. Although fixed kernel utilization distributions give a better estimate of the intensity of area use than MCPs (Seaman et al. 1998), in this particular case fixed kernel 99% utilization distributions were often fragmented and excluded area that sheep were logically required to have passed through in the 3-4 hours between relocations when moving from one portion of their home ranges to another.
The number of sorties launched each day into MOA1 and MOA2 provided an estimate of area-wide overflight intensity and were obtained from USAF personnel. The CL study area lies entirely with MOA1 while the WP study area is located in both MOA1 and MOA2 (Chapter 3, Fig. 3.1). Because the number of daily sorties in MOA1 was highly correlated with the daily total of sorties in MOA1 and MOA2 combined (r2=0.97), we used the number of sorties in MOA1 as our estimate of relative intensity of flying on a daily basis for both the CL and WP study areas. For each sheep, we averaged the number of sorties launched for the actual days that a sheep generated GPS locations within each 2-week sequential period.
We assessed the effect of year and sequential 2-week period on the number of sorties launched per day with ANOVA (PROC GLM, SAS V8e Windows).
We used multiple analysis of covariance (MANCOVA; PROC GLM, SAS V8e Windows) to test whether or not the average daily number of sorties in MOA1 within each 2-week period increased the precision of our estimates of the effect of the study area, and the nested effects of years within study areas and sequential periods within years within study areas (class variables) on dependent behavioral variables. The vector of behavioral data included: 1) 2-week MCP home range size (km2) for each sheep, and 2) sum of daily distances moved (km) within the 2-week home range for each sheep.
We also used the sorties covariate in a separate multiple analysis of covariance of the influence of the class variables on the vector of habitat use within the 2-week home range for each sheep. The vector of habitat use for each sheep was composed of 11 elements that included the proportional use of three landcover classes (timber, grass-shrub-herb, and sparsely vegetated; unknown landcover was used to calculate proportions but excluded from further analysis), proportional use of four aspects (90o arcs including N, S, E, and W), average elevation, average slope, average terrain ruggedness, and median maximum vegetation greenness (NDVI), all estimated from the 50-120 GPS relocations of each sheep in each 2-week sequential period. Class variables were fixed effects and expected to be significant sources of variation in the dependent variables due to different habitats between study areas, different annual conditions among years, and the natural phenological progression of forage plants and energetic status of sheep across the eight 2-week periods within years. We assessed whether the average number of daily sorties was related to the dependent variables and compared Type I SS and Type III SS for the class effects to estimate whether the number of sorties influenced the significance of class effects. All analyses were conducted twice; once with the covariate entered first and once with the covariate entered last.
There was some multi-collinearity in our data. Sparsely-vegetated areas tended to be less green and at higher elevation and timbered areas tended to be more green and at lower elevation. There were also correlations between East and West aspects, and East aspects tended to have less steep slopes. To assess the influence of multi-collinearity, we deleted landcover class and East aspects from our data, reanalyzed, and compared the conclusions to those reached from consideration of all data.
Overall, 523 observations were available for the behavioral analyses and 567 observations were available for the habitat use analyses. These were distributed within the two-study-area by four-year by eight-2-week-period design for an average of 8-9 sheep per cell. All statistical tests were conducted at the a priori α=0.05 level of significance.
We used simulation modeling to assess the power of our analyses to detect significant differences in the multiple analyses of covariance (Appendix F).
Potential influence of military aircraft sorties on behavior and habitat use
There were no differences in conclusions reached due to inclusion/exclusion of correlated independent variables.
From early April – early August, the average number of sorties launched per day was significantly (P < 0.001) different among years (22.9, 12.5, 25.7 and 31.8 sorties/day for 1999-2002, respectively) and among sequential periods within years (P < 0.001; 21.8, 21.3, 12.7, 8.0, 28.7, 12.8, 52.3 and 18.2 sorties/day for sequential periods from late-April through early-August, respectively) (Fig. 4.1). Sorties were at their lowest annual values during and immediately after lambing (late-May and early-June) and were highest in late-July. Spikes in sorties accompanied actual Cope Thunder Military Flying Exercises (MFEs) in early- May, late-June and late-July (Fig. 4.1). The average number of sorties launched per day was significantly greater (P < 0.001) during actual MFEs (59.8) than during periods without MFEs (18.7).
 |
| Figure 4.1. Average number of military sorties launched per day into MOA 1, 1999-2002, in eight 2-week sequential periods within years. |
There were overall significant (P < 0.001) effects of study area, year within study area, and sequential period within years within study areas on home range size and distance moved. The full model explained 42% of the variance in minimum total distance traveled during the 2-week periods and 62% of the variance in 2-week MCP home range size. When considered alone (Type I SS) and entered first in the modeling process, the average number of sorties/day was significant (P < 0.001) for both distance moved and home range size. However, the sorties covariate was 1) never significant (distance P = 0.953; home range size P = 0.14) once the effects of study area, year within study area, and sequential period within year within study area were in the model (Type III SS), and 2) did not change the significance of any of the main class effects.
There were overall significant (P < 0.001) effects of study area, year within study area, and sequential period within years within study areas on the vector of habitat use (proportional use of landcover classes (3), proportional use of aspects (4), average slope, average elevation, average terrain ruggedness, and median maximum vegetation greenness (NDVI)). The full model explained 27-57% of the variance in individual elements of the habitat use vector. When considered alone (Type I SS) and entered first in the modeling process, the average number of sorties/day was significant (P < 0.0008) for proportional use of grass-shrub-herb landcover, proportional use of north and south aspects, and average terrain ruggedness. However, the sorties covariate was never significant (P > 0.084) once the effects of study area, year within study area, and sequential period within year within study area were in the model (Type III SS) and never changed the significance of any of the main class effects.
There was a visual suggestion that average home range size (Fig. 4.2), 2-week travel distance (Fig. 4.3) and terrain ruggedness used (Fig. 4.4) may have been increasing from 10 through 30 sorties per day. However, separate analyses of the relationships between the dependent variables and the number of sorties per day within this restricted range did not show a significant (P > 0.405) relationship. There was no visual suggestion of a relationship between sortie/day and any other habitat use variable.
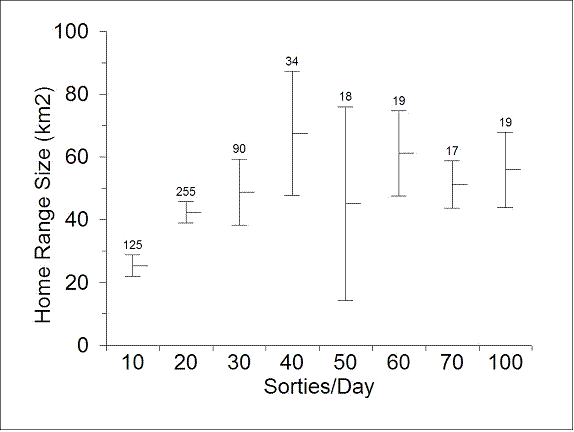 |
| Figure 4.2. Average MCP home range size of sheep in relation to number of military sorties launched per day into MOA 1, interior Alaska, late-April through early-August, 1999-2002. Bars encompass the mean plus or minus 2 standard errors. Labels are the number of sheep*year*sequence combinations that yielded the mean. |
 |
| Figure 4.3. Average 2-week minimum total distance traveled by sheep in relation to number of military sorties launched per day into MOA 1, interior Alaska, late-April through early-August, 1999-2002. Bars encompass the mean plus or minus 2 standard errors. Labels are the number of sheep*year*sequence combinations that yielded the mean. |
 |
| Figure 4.4. Average terrain ruggedness used by sheep in relation to number of military sorties launched per day into MOA 1, interior Alaska, late-April through early-August, 1999-2002. Bars encompass the mean plus or minus 2 standard errors. Labels are the number of sheep*year*sequence combinations that yielded the mean. |
Our simulation assessment of the statistical power of our tests (Appendix F) suggested that if the magnitude of mean differences in behavior and habitat use observed in Figures 4.2-4.4 were real, it would have taken a sample size of approximately 20 sheep per study area to declare them statistically significant.
Behavior and habitat use by study area, year and sequence
Average home range size in West Point (47.8 km2) was significantly (P < 0.0001) larger than average home range size in Cirque Lakes (14.0 km2) and there was significant ( P < 0.0001) variation in home ranges sizes among years within study areas and among sequential periods within years within study areas at West Point (Fig. 4.5) and Cirque Lakes (Fig. 4.6). Particularly for West Point (Fig. 4.5), home range sizes became quite large in late-June, about a month after lambs were born, and remained large thereafter through early August. Increase in home range size from late-June onward was evident for Cirque Lakes only in 1999.
Average minimum total distance traveled within 2-week sequential periods at West Point (51.3 km) was significantly (P < 0.0001) larger than distance traveled at Cirque Lakes (35.8 km) and there was significant (P < 0.0001) variation in distance travelled among years within study areas and among sequential periods within years within study areas at West Point (Fig. 4.7) and Cirque Lakes (Fig. 4.8). There was a tendency toward greater travel distance as the seasons progressed in both study areas (Figs. 4.7, 4.8). Total distance traveled was quite high for West Point in 2001 in all periods except early- and late-June (Fig. 4.7), and distance traveled was highest in Cirque Lakes during late-June and early-July of 1999 (Fig. 4.8).
 |
| Figure 4.5. Average MCP home range size (km2) for Dall’s sheep in the West Point study area, interior Alaska, late-April through early-August, 1999-2002. Labels indicate presence (1) or absence (0) of major military flying events (MFE). |
 |
| Figure 4.6. Average MCP home range size (km2) for Dall’s sheep in the Cirque Lakes study area, interior Alaska, late-April through early-August, 1999-2002. Labels indicate presence (1) or absence (0) of major military flying events (MFE). |
 |
| Figure 4.7. Average minimum total distance (km) traveled in 2-week sequential periods for Dall’s sheep in the West Point study area, interior Alaska, late-April through early-August, 1999-2002. Labels indicate presence (1) or absence (0) of major military flying events (MFE). |
 |
| Figure 4.8. Average minimum total distance (km) traveled in 2-week sequential periods for Dall’s sheep in the Cirque Lakes study area, interior Alaska, late-April through early-August, 1999-2002. Labels indicate presence (1) or absence (0) of major military flying events (MFE). |
In both study areas, use of the sparsely vegetated landcover class accounted for more than half of the usage and there was slightly greater (P < 0.0001) proportional use of the sparse landcover class in West Point (0.606) compared to Cirque Lakes (0.539; Figs. 4.9, 4.10). Some exceptions to this were evident for West Point in early-April through early-May in 1999 and late-May through early-June in 2001 (Fig. 4.9) when use of sparsely vegetated areas was reduced well below half of all landcover usage. Sparsely vegetated areas tended to be at higher elevation.
There was significantly (P < 0.0001) less proportional use of the timber landcover class in West Point (0.057, Fig. 4.11) compared to Cirque Lakes (0.187, Fig. 4.12). Substantial use of the timber class at Cirque Lakes was particularly evident from early-April through early-July in 2001, and from late-June through early-August in 2002 (Fig. 4.12). Much of the use of timber in Cirque Lakes was associated with sheep making substantial use of the bluffs of the Charley River in those years and sequential periods.
Conversely, there was significantly (P < 0.0001) more proportional use of the grass-shrub-herb landcover class in West Point (0.314, Fig. 4.13) compared to Cirque Lakes (0.126, Fig. 4.14). In both study areas, there was a tendency toward increased relative use of the grass-shrub-herb landcover class from early-April through early-June compared to the remainder of the sequential periods (Figs. 4.13, 4.14) but this was not evident in all years.
 |
| Figure 4.9. Average proportional use of the sparsely vegetated landcover class for Dall’s sheep in the West Point study area, interior Alaska, late-April through early-August, 1999-2002. Labels indicate presence (1) or absence (0) of major military flying events (MFE). |
 |
| Figure 4.10. Average proportional use of the sparsely vegetated landcover class for Dall’s sheep in the Cirque Lakes study area, interior Alaska, late-April through early-August, 1999-2002. Labels indicate presence (1) or absence (0) of major military flying events (MFE). |
 |
| Figure 4.11. Average proportional use of the timber landcover class for Dall’s sheep in the West Point study area, interior Alaska, late-April through early-August, 1999-2002. Labels indicate presence (1) or absence (0) of major military flying events (MFE). |
 |
| Figure 4.12. Average proportional use of the timber landcover class for Dall’s sheep in the Cirque Lakes study area, interior Alaska, late-April through early-August, 1999-2002. Labels indicate presence (1) or absence (0) of major military flying events (MFE). |
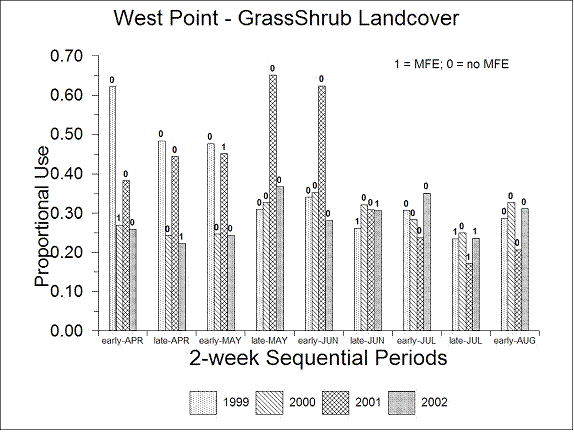 |
| Figure 4.13. Average proportional use of the grass-shrub-herb landcover class for Dall’s sheep in the West Point study area, interior Alaska, late-April through early-August, 1999-2002. Labels indicate presence (1) or absence (0) of major military flying events (MFE). |
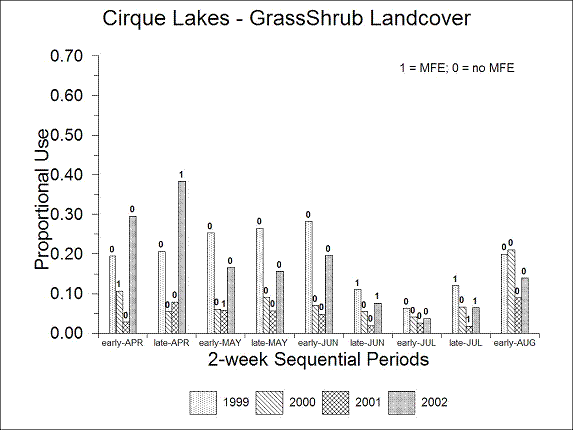 |
| Figure 4.14. Average proportional use of the grass-shrub-herb landcover class for Dall’s sheep in the Cirque Lakes study area, interior Alaska, late-April through early-August, 1999-2002. Labels indicate presence (1) or absence (0) of major military flying events (MFE). |
From early-April through early-May in 1999 and from early-April through early-June in 2001, sheep at West Point used sites that would ultimately have relatively high plant biomass at peak vegetation development (Fig. 4.15). This pattern of use was coincident with increased use of the grass-shrub-herb landcover class in the same sequential periods and years (Fig. 4.13). The average NDVI at used sites for Cirque Lakes (0.101; Fig. 4.16) was slightly greater (P < 0.001) than at West Point (0.079) but this may have been influenced by the greater use of the timber landcover class at Cirque Lakes (Fig. 4.12) compared to West Point (Fig. 4.11). NDVI estimates greenness in the vegetation canopy, and if the canopy is unavailable to sheep (i.e. the tops of trees), it may not indicate greater forage availability.
Proportional use of elevation (Figs. 4.17, 4.19) and slope (Figs. 4.18, 4.20) at West Point was more consistent among years and sequential periods than at Cirque Lakes but, on average, sheep used slightly higher elevations (1,346m vs. 1,195m; P < 0.0001) at West Point compared to Cirque Lakes, and slightly steeper slopes (51.8% vs. 47.1%; P <0.0001) at Cirque Lakes compared to West Point, respectively.
At West Point, used terrain was slightly less rugged than at Cirque Lakes (71.3 vs. 75.4, respectively, P < 0.0001) and this difference was most pronounced from early-April through early-May (Figs. 4.21, 4.22). Terrain shadow and unknown landcover classes at sheep locations tended to be more prevalent in Cirque Lakes (14.8%) than in West Point (2.9%). This probably reflected steeper and more rugged terrain in Cirque Lakes which would be more likely to be in shadow on a satellite image. Thus, we probably underestimated the difference in use of rugged terrain between Cirque Lakes and West Point.
 |
| Figure 4.15. Average maximum relative green plant biomass (NDVI) at sites used by Dall’s sheep in the West Point study area, interior Alaska, late-April through early-August, 1999-2002. Labels indicate presence (1) or absence (0) of major military flying events (MFE). NDVI estimates obtained from a single TM image from 11 July 1994 near the peak of vegetation development for that year. |
 |
| Figure 4.16. Average maximum relative green plant biomass (NDVI) at sites used by Dall’s sheep in the Cirque Lakes study area, interior Alaska, late-April through early-August, 1999-2002. Labels indicate presence (1) or absence (0) of major military flying events (MFE). NDVI estimates obtained from a single TM image from 11 July 1994 near the peak of vegetation development for that year. |
 |
| Figure 4.17. Average elevation use for Dall’s sheep in the West Point study area, interior Alaska, late-April through early-August, 1999-2002. Labels indicate presence (1) or absence (0) of major military flying events (MFE). |
 |
| Figure 4.18. Average elevation use for Dall’s sheep in the Cirque Lakes study area, interior Alaska, late-April through early-August, 1999-2002. Labels indicate presence (1) or absence (0) of major military flying events (MFE). |
 |
| Figure 4.19. Average slope (%) use for Dall’s sheep in the West Point study area, interior Alaska, late-April through early-August, 1999-2002. Labels indicate presence (1) or absence (0) of major military flying events (MFE). |
 |
| Figure 4.20. Average slope (%) use for Dall’s sheep in the Cirque Lakes study area, interior Alaska, late-April through early-August, 1999-2002. Labels indicate presence (1) or absence (0) of major military flying events (MFE). |
 |
| Figure 4.21. Average terrain ruggedness use for Dall’s sheep in the West Point study area, interior Alaska, late-April through early-August, 1999-2002. Labels indicate presence (1) or absence (0) of major military flying events (MFE). |
 |
| Figure 4.22. Average terrain ruggedness use for Dall’s sheep in the Cirque Lakes study area, interior Alaska, late-April through early-August, 1999-2002. Labels indicate presence (1) or absence (0) of major military flying events (MFE). |
Proportional use of aspect differed (P < 0.0001) between study areas for all aspects. Sheep at both West Point (Figs. 4.23-4.26) and Cirque Lakes (Figs. 4.27-4.30) spent the greatest proportion of their time on south (0.37 and 0.43, respectively) and west (0.36 and 0.30, respectively) aspects. When sheep decreased their use of south and west aspects from late-June through early August at West Point (Figs. 4.23, 4.24) and Cirque Lakes (Figs. 4.27, 4.28) they concurrently increased their use of north and east aspects in both study areas (West Point , Figs. 4.25, 4.26; Cirque Lakes Figs. 4.29, 4.30). These shifts in proportional use of aspect were less pronounced at Cirque Lakes (Figs. 4.27-4.30) than at West Point (Figs. 4.23-4.26).
 |
| Figure 4.23. Average proportional use of southern aspects (136-225o) for Dall’s sheep in the West Point study area, interior Alaska, late-April through early-August, 1999-2002. Labels indicate presence (1) or absence (0) of major military flying events (MFE). |
 |
| Figure 4.24. Average proportional use of western aspects (226-315o) for Dall’s sheep in the West Point study area, interior Alaska, late-April through early-August, 1999-2002. Labels indicate presence (1) or absence (0) of major military flying events (MFE). |
 |
| Figure 4.25. Average proportional use of northern aspects (316-045o) for Dall’s sheep in the West Point study area, interior Alaska, late-April through early-August, 1999-2002. Labels indicate presence (1) or absence (0) of major military flying events (MFE). |
 |
| Figure 4.26. Average proportional use of eastern aspects (046-135o) for Dall’s sheep in the West Point study area, interior Alaska, late-April through early-August, 1999-2002. Labels indicate presence (1) or absence (0) of major military flying events (MFE). |
 |
| Figure 4.27. Average proportional use of southern aspects (136-225o) for Dall’s sheep in the Cirque Lakes study area, interior Alaska, late-April through early-August, 1999-2002. Labels indicate presence (1) or absence (0) of major military flying events (MFE). |
 |
| Figure 4.28. Average proportional use of western aspects (226-315o) for Dall’s sheep in the Cirque Lakes study area, interior Alaska, late-April through early-August, 1999-2002. Labels indicate presence (1) or absence (0) of major military flying events (MFE). |
 |
| Figure 4.29. Average proportional use of northern aspects (316-045o) for Dall’s sheep in the Cirque Lakes study area, interior Alaska, late-April through early-August, 1999-2002. Labels indicate presence (1) or absence (0) of major military flying events (MFE). |
 |
| Figure 4.30. Average proportional use of eastern aspects (46-135o) for Dall’s sheep in the Cirque Lakes study area, interior Alaska, late-April through early-August, 1999-2002. Labels indicate presence (1) or absence (0) of major military flying events (MFE). |
This portion of the study addressed sheep response at the within-home-range scale and the sampling units were the GPS collared sheep which were assumed to represent a random sample of the adult female sheep in the study area.
We observed no significant effect of the number of military aircraft sorties on 2-week within-home-range scale behavior (total distance moved, home range size) or habitat use (proportional use of landcover classes and aspect classes, average elevation, slope, terrain ruggedness, and vegetation biomass) that exceeded natural variation due to study area, year, and season. There was substantial variance in behavior and habitat use between study areas, between years within study areas, and among 2-week sequential periods within years within study areas. Once this variance was accounted for, the number of military aircraft sorties added no further explanatory power to the assessment of variance in sheep behavior and habitat use. Considering the small proportion of variance in the activity and foraging efficiency of sheep (<1%) that was explained by the number of military aircraft observed during field observation sessions (Chapter 3, this document), it is not surprising that we detected no effect of the number of military flight sorties on sheep behavior or habitat use at the larger, within-home-range, scale of analysis.
Because sheep in the two study areas had been exposed to military overflights for at least 21 years prior to the initiation of this study, sheep remaining in the study areas may have become acclimated to military overflights. If such acclimation had occurred, then the likelihood of detecting a significant response of sheep to military aircraft sortie levels would be diminished. Because no pre-treatment (i.e., observations without a history of military overflights) data are available, it is impossible to assess whether sheep populations and sheep performance (productivity, behavior or habitat use) have acclimated to the history of military overflights in the two study areas. However, we do note that sheep have persisted during the 22 years between establishment of the MOA’s and the beginning of these studies, and remained relatively stable in numbers during the past 5 years, in both study areas which have a relatively long history of use as MOAs. Relatively low sorties/day during lambing and post-lambing (late-May and early-June; Fig. 4.1) may have minimized any potential effects of overflights on sheep.
We emphasize that the studies conducted here reflect an average background level of slightly less than 20 sorties/day which increased 3-fold to an average of about 60 sorties/day during MFEs, and that peaked at about 95 sorties/day during one year-period combination (Fig. 4.1). If average background sortie levels, average MFE sortie levels, peak sortie levels and frequency, or the seasonal pattern of sorties (Fig. 4.1) changes substantially from the values observed in this study, then the conclusions reached here regarding a detectable effect may not be applicable.
The nature of the implementation of the Cope Thunder Military Flying Exercises (MFE) prevented a direct analysis of the effects of the MFEs or the mitigation associated with the Cirque Lakes study area on within-home-range scale response of sheep. Not all scheduled MFEs were conducted, mitigation was not continuous at Cirque Lakes during implemented MFEs, and substantial numbers of sorties were launched in MOA1 and MOA2 when MFEs were not occurring. It was impossible to contrast implemented/not implemented MFEs among years while controlling for season (e.g., there was only one June in each year, an MFE was conducted or not, and implementation or not of MFEs was not balanced among years and seasons). The design of the study described here was essentially correlational because there was no control of the number of sorties launched and there were no sequential periods without sorties.
The power of the analyses were limited to some degree (Appendix F) due to relatively small number of sheep that were collared in each of the study areas that turned out to be quite different in characteristics and in behavior and habitat use by sheep. However, we did detect statistically significant differences in use of elevation and sparsely vegetated areas between the two study areas that were only ~11% of the larger value in the comparison.
Approximately 20 sheep per study area (twice as many as available) would have been required to declare the observed mean differences in behavior by sortie class (Figs. 4.2-4.3) to be statistically significant. These mean values (Figs 4.2-4.3) suggest that home range size and distance moved within 2-week periods were quite low for the lowest sortie levels (10/sorties/day) compared to sortie levels of 20/day and greater.
Even if there had been sufficient statistical power to declare these differences (Figs. 4.2, 4.3) significant, it is not clear that the tendency toward increased home range size and distance moved for sortie levels > 10/day could have been clearly attributed to military overflights. The lowest sortie levels occurred primarily during the last trimester of pregnancy, lambing, and immediately after lambing when home range size and movement rates might naturally be expected to be low as a result of restrictions on movement of the adults by neonates.
Clear resolution of the potential effects of military aircraft sortie level on behavior and habitat use of sheep at the home range scale will require 1) replicated study areas, years, and seasons that differ only in the number of military sorties, 2) an adequate sample size of sheep (>20 collared sheep/study area), and 3) experimental control of the number of sorties that sheep are exposed to, including replicates with no sorties and replicates with no history of sorties. It is extremely unlikely that the first requirement can ever be satisfied due to the natural variance in areas, years, and seasons. The results of this study do suggest that, under the constraints of relatively few marked sheep and a system with a relatively long history of exposure to military overflights, any effects of major flying exercises on sheep behavior and habitat use were not more dramatic than the natural variance in behavior and habitat use that would be expected to result from variance among study areas, years, and seasons within years.
We assessed the effect of the number of military sorties launched on home range size, movement rates, and use of habitats for Dall’s sheep in interior Alaska over four years, 8 sequential 2-week-long periods within years, and two study areas. The average number of sorties launched per day was greater (59.8 sorties/day) during major flying exercises than during routine operational days (18.7 sorties/day). There was significant variation in sorties/day among years and among sequential periods within years. There was significant variation in home range size, movement rates, and habitat use between study area, among years within study areas, and among periods within years within study areas. The effect of sorties/day was never a significant source of variation in home range size, distance traveled, or habitat use, once the effects of study area, year, and period were in statistical models. The lack of demonstrated significant response by sheep to sorties/day in this study may have been due to: 1) relatively rare exposure of sheep to low, close, and loud military overflights and relatively small-magnitude behavioral response to these overflights (see Chapter 3); 2) a long period (22 years prior to study initiation) of acclimation by sheep to military overflights in the area; 3) inability to detect an effect of routine background sorties/day as we had no study area without military overflights for comparison; or 4) insufficient statistical power to detect small-magnitude effects that was a result of having a relatively small number of radio-collared sheep.
GENERAL CONCLUSIONS and MANAGEMENT IMPLICATIONS
We conclude that the levels of military activity which we observed in this study during Major Flying Exercises (~60 sorties/day) generally did not cause statistically significant effects on sheep behavior and habitat use at the home range scale when compared to the background level of military sorties (~ 20 sorties/day). When differences in feeding efficiency and activity of groups of sheep were observed, military aircraft activity accounted for a small proportion of the total variance in sheep behavior, the direction of these effects were variable, and there were no overt indications that military overflights affected sheep populations during 1999-2002. Few of the overflights that we observed were low or loud (7% of observed military overflights were below 1500 m and within 1.6 km of observed sheep) and the mitigation measures in place at the Cirque Lake study site provided no detectable reduction in sheep response to military overflights. We base these conclusions on a number of findings. Dall’s sheep population in Yukon-Charley Rivers National Preserve were stable from 1997-2002. We found no differences in Dall’s sheep population trends in an area mitigated for low-level military overflight verses an area that was not that could be attributable to low-level military jets. No statistical differences were found in Dall’s sheep pregnancy rates, lamb to ewe ratios, yearling to ewe ratios or survival in an area mitigated for military overflights verses an area that was not. In fact, ewe body weights were higher at a non-mitigated study site verses a mitigated site. The number of overflights we observed over Dall’s sheep on a given day did not influence Dall’s sheep behavior. Significant differences occurred in the feeding efficiency of Dall’s sheep when behavior was examined in the 10 minutes before, during and 10 minutes after an overflight event. In models that included all seasons, Dall’s sheep ewe feeding efficiency was higher after the overflight event in comparison to before the overflight event. In models that just examined lambing season and early summer (May, June and July), sheep feeding efficiency was higher before the overflight event in comparison to during the event. In models that considered the proximity and sound level of overflight events, a higher proportion of sheep were active with increasing sound levels. In models that just examined lambing season and early summer, more sheep were active with closer military overflights. Factors other than military overflights explained most of the variability in the observed behavior of sheep. Of particular note in this regard was the time of year (date sequence nested within year and study site). Also of note is the rarity of low (20 overflights <1500 m) and loud (4 overflight events with 2-min mean levels > 71dBA) military overflight events during 2 years of field work and the lack of correlation between Cope Thunders (a large scale military flying exercise) and the number of military aircraft seen by observers on the ground. Because low and loud aircraft were rare we were not able to test the effectiveness of the 5,000 foot AGL military aircraft mitigation measure at the Cirque Lake study area. Dall’s sheep home ranges sizes, distances moved, and habitat use did not vary due to the number of military sorties launched per day.
Although we found little evidence of effects of military overflights on sheep, our data do suggest a number of management considerations:
1) Sheep moved between study areas - Considerable variation existed in number of observed sheep at individual survey units from year to year yet overall sheep numbers in Yukon-Tanana Uplands were relatively stable. This may have been due to movement of sheep from one survey unit to another as we observed for our radiocollared sheep. Management of sheep in this area therefore, should be directed at the entire Yukon-Tanana Uplands and not at individual patches of sheep habitat. In addition, protection of sheep in the Yukon-Tanana Uplands needs to consider travel routes to suitable patches of habitat as well as the habitat patches themselves. Movement of sheep between disjunct patches of habitat could help ameliorate the affects of unfavorable conditions experienced at one habitat patch. Because travel routes are important for interchange among subpopulations (Geist 1971, Nichols and Bunnel 1999) disruption of travel routes could fragment this sheep population. Small isolated populations of sheep would be much more vulnerable to a variety of factors than would a larger contiguous sheep population.
2) Small trend areas are not directly appropriate for assessing sheep performance in larger regions - Variation in number of observed sheep in individual survey units from year to year yet stability in the overall sheep numbers in Yukon-Tanana Uplands suggests that small “trend areas” are not appropriate for determining trends in sheep populations in the Yukon-Tanana Uplands. To be most useful, surveys need to cover large areas including numerous patches of sheep habitat.
3) Small sub-populations are quite variable and likely sensitive to stochastic events - We observed substantial annual and study site variation in Dall’s sheep pregnancy rates, lamb:100 ewes, and yearlings:100 ewes. Short-term or localized conditions therefore may have substantial effects on the local reproductive success of Dall’s sheep and indicate that Dall’s sheep subpopulations may be sensitive indicators of small scale local environmental conditions. This sensitivity needs to be considered when long-term changes in the environment are contemplated in the Yukon-Tanana Uplands. Changes in the nature of military flying activity in interior Alaska MOAs would therefore, need to be evaluated to determine how sheep populations may respond.
4) Military overflights were less than expected and quite variable. We observed considerable fluctuations in the extent of military flying activity from one year to the next, from one date sequence to the next, and from one Cope Thunder Exercise to the next. During the 4 years of this study, scheduled flying activities were curtailed for a number of reasons. We do not know how typical the level of flying activity we observed is for the interior Alaska MOAs. We do know that we did not evaluate the effects of military aircraft overflights levels presented as typical in the Final Environmental Impact Statement, Alaska Military Operations Areas (Department of the Air Force 1995).
5) Loud and low military jet overflights were rare events during this study. Loud and low jets can affect Dall’s sheep behavior. Reduction of the potential for low and loud jets would therefore reduce the potential of affecting Dall’s sheep behavior. Altitude floors, such as the 5,000 m floor associated with the current mitigation measure, is therefore an appropriate means by which to reduce potential impacts to Dall’s sheep. Anecdotal observations indicate that multiple jets produced greater sound levels than did single aircraft. Therefore, a second possibility might be to restrict the number of aircraft flying together.
2004 Sheep Report
http://www.nps.gov/yuch/Expanded/key_resources/sheep/sheep_2004.htm
Doug Beckstead
December 8, 2004