![]()
![]()
![]()
by
J.G. Pope
M.A.F.F. Fisheries Laboratory
Lowestoft
1. INTRODUCTION
Limiting fishing mortality by catch quotas (total allowable catches or TACs) requires the input of scientific advice if it is to be successful. In some regions, aspects of this advice are fairly routine, while other aspects are in clear need of development. In other regions, almost everything remains to be developed if this method of managing fisheries is to be adopted. This paper seeks to review briefly the scientific methods used and their limitations. It seeks to indicate where new technique is needed and where more dialogue is required between scientists, administrators and fishermen.
2. OBJECTIVES OF MANAGMENT
2.1 True objectives and proximate goals
The management of fisheries may be aimed at a number of possible objectives. For example, it might seek to maximize the catch of the fishery, to maximize the employment of fishermen and ancilliary workers, to maximize the economic yield either to vessel owners, fishermen or government. Again it might seek to maximize foreign earnings or ensure a regular supply to consumers. There thus tend to be a bewildering complexity of possible objectives, many of which are conflicting and few of which are properly quantified. For these reasons the aims of management are often expressed as the attainment of some proximate goal such as (a) to maintain the current level of fishing mortality or some proportion of it, and (b) to attain some specified level of fishing mortality such as F max which gives the maximum sustainable yield or F0.1 which approximates to the maximum economic yield.
Clearly these proximate goals are aimed towards the yield-maximization or profit-maximization objectives of fisheries management but the extent that they will meet this latter objective or indeed any other objective is not very clear. A detailed study of the relationship of the proximate goals to the objectives of management would be very useful. Has it been made anywhere?
I would expect that the proximate goals would prove valuable as a way of approaching some objectives, counter-productive for other objectives and irrelevant to some other objectives. Given the proximate goals, however, catch quotas are an obvious means of obtaining them.
2.2 Problems with the definition of proximate goals of fisheries management
Although the proximate aims of fisheries management give the impression of crispness, there are in fact several points in the definition that need clearing up. How accurately do they need to be attained? Would a systematic bias of 10 percent either way worry anyone? Would they need to be precise each year or would it be sufficient if the goal was met on average? Do we need to specify a maximum variability? Where the link between fishing effort and fishing mortality is fixed the answer is probably that the variability of the goal might well have important economic costs. Both Rivard (1981) and Pope (1981) consider that TAC management should have a 10-percent coefficient of variation, if this is achievable. Where the link is variable, as it often is for pelagic species, very precise TACs may be inappropriate. Certainly then there is a need for discussion on how accurately and precisely proximate goals need to be set, between the scientists who have to estimate TACs, the administrators who apportion them and the fishermen who suffer their consequencies. The answer to scientific problems, such as how precise various data sets really need to be, will ultimately come from the answer to this question. So will the answer to the question of what stocks might reasonably be managed by TAC.
Another problem with the proximate aims is just what is meant by fishing mortality. Fishing mortality (the instantaneous coefficient of fishing mortality) occurs as a potentially-independent variable on each separate age of fish. Clearly some overall measure of fishing mortality is required and the quick answer is usually that the mortality referred to is the fully-recruited mortality. What however is this?
The text table shows several different sets of fishing mortality at age:
Fishing Mortality at Age
| A G E | ||||||||
|---|---|---|---|---|---|---|---|---|
| Set | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
| I | 0 | .4 | .8 | .8 | .8 | .8 | .8 | .8 |
| II | .4 | .8 | .8 | .8 | .8 | .8 | .8 | .8 |
| III | .8 | .8 | .8 | .8 | .8 | .8 | .8 | .8 |
For sets I, II and III, the fully-recruited fishing mortality is 0.8 but obviously set III has more mortality than set II, which in turn has more than set I. If these results came from three successive years then we would hardly be justified in assuming that the mortality had remained the same. The goal that the mortality should stay the same as last year's thus should perhaps be that the fully-recruited mortality should remain the same and the exploitation pattern (the proportions in which this is distributed over the various ages) should also remain unchanged. The problem is that the exploitation pattern can be observed to vary considerably from year to year. The problem then is to express the result of all mortalities as a single figure. Shepherd (1982) gives two estimators of overall fishing mortality which go some way towards this need. Both are weighted means of fishing mortality at age. The first (Fc) is weighted by the catch number at age that would result from a single cohort of fish with these mortalities at age. Under this measure, run II would show the same result as run I. In other words, it might be thought of as being generated with the same level of fishing effort but with a smaller mesh size. It would thus be a useful measure where a mesh change was applied. The second means (Fp) is weighted by the population number at age. It would thus indicate that mortality was higher for set II than set I. It would be useful as a general measure of biomass erosion.
Another approach is simply to assume a constant exploitation pattern and to impose this on the interpretation of fishing mortality. This is the approach adopted in many least-squared interpretations of catch at age data. For example, that of Pope and Shepherd (1982) expresses fishing mortality on age j in year i as Fi × Sj, where Fi is an annual fishing mortality and Sj the exploitation pattern. Such an interpretation does therefore give a single estimation of mortality for each year. In some senses this is justified because fisheries scientists need the concept of a fixed exploitation pattern in order both to estimate Fmax levels and to forecast catch levels (e.g. to estimate TACs). Thus there may be some justification in finding what is in a sense the best average exploitation pattern and using it. In another sense, however, it is only reasonable if departures from this separability model are random since systematic departures (e.g. a trend with time) will cause bias. Clearly the fishing mortalities on the several ages of fish cannot always be adequately described by a single dimensional vector. For example, a principal component analysis carried out on the North Sea haddock from 1961 to 1976 indicated that there were in fact two almost orthogonal components where fishing mortality at age was considered as variables and years as cases. These indicated that the mortality on younger ages formed one component (corresponding to by-catches in the industrial fishery), while the mortality on the older ages formed another component. These two components appeared to account for most of the coherent variation (63 percent) of fishing mortality at age in these years and properly, therefore, we should have to talk of the fishing mortality on haddock in terms of the level of each of these two components. At Lowestoft we are currently developing a two-fleet version of the Pope and Shepherd (1982) method which should enable this to be carried out in a systematic fashion. Given such interpretations, the proximate goal of management might be to maintain the mortality on both components at their current levels, or perhaps to increase one by 10 percent and decrease one by 10 percent. This would, of course, mean setting separate TACs for separate fleet segments. It underlines what is an often neglected problem: that the proper level of a TAC depends to some extent on its allocation to different fleets or gear types or countries.
2.3 Timing of Advice
The advice fisheries scientists can give on future fish catches and hence TACs is very much a function of how far ahead the advice has to be given. The shorter the time between the advice being given and being used, the more precise and accurate it is likely to be. As the time extends, changes in the recruitment levels of new year classes of fish, unknown catch levels in intermediate years and changes in exploitation pattern all add sources of error to the prediction. Thus in principle administrators need to think of ways of shortening the time between advice being given and being used. This may perhaps be achieved by TACs being set provisionally and then being adjusted, if this is necessary, nearer to the time of application in the light of fresh evidence. This problem of timing is a very serious problem for fisheries scientists which administrators should perhaps be more aware of.
3. ESTIMATION OF TACs
The estimation of appropriate TACs is usually undertaken by fisheries scientists. The methods used for estimation vary according to the particular problems associated with the fishery and with the availability of data.
3.1 Variability of TACs
Scientific advice for fisheries management by TAC is often given so as to satisfy one of the proximate goals. If these goals are to be satisfied accurately then detailed knowledge of the biomass of fish by age or size is needed. Since variation in recruitment to the fishery causes this to vary with time, it also follows that TACs will generally need to be re-assessed with time, often on an annual basis. The effect of variation in recruitment on TACs is important to understand because it may determine the types of situation for which TAC management is applicable.
Fish catches are often composed of a number of ages of fish. Indeed where this is not the case, the prediction of a future catch (such as a TAC) may be very problematic. The yield in year t - Y(t) - may be written simply in terms of the numbers of fish recruiting each year R(i), the weight of age j - W(j), the exploitation proportion Ex (t,j) for each age j in the current year t and the proportion of the recruitment surviving from year i to year t - SU(t,i).
Thus,
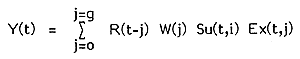 | (1) |
where, g is the oldest exploited age.
Clearly changes in any of these factors from year to year can change the catch. In practice all do change to some extent from year to year. Table 1 shows the range of variation of recruitment for some North Sea fish stocks over a number of years. In higher latitudes this often proves the greatest source of variation in yield. This may of course not be the case for fisheries in other climatic regions. For example, the rather systematic declines of biomass seen in the groundfish-survey results from the Gulf of Thailand; Pope (1979) suggests that there the recruitment variation may be less, although systematic changes are quite possible. Since the objective of the TAC management is often to stabilze levels of fishing mortality and hence the Su(t,i) and Ex(t,j) terms in equation (1), we may conveniently regard yields as weighted sums of recruitment data. The weighting factor is of course:
W(j) Su(t,i) Ex(t,j)
Following equation 6.31 of Beverton and Holt (1957) we may express this simply as:
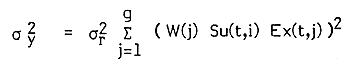 | (2) |
where, σ2y and σ2r are the variances of yield and recruitment, respectively.
Table 1
Recruitment variations in North Sea fisheries, 1963–1978
Number adjusted to a value of 100 for the most outstanding year-class of each species
Ratio is the ratio between the numbers in the strongest
and weakest year-classes in the period
| Year | Cod | Haddock | Whiting | Saithe | Plaice | Sole | Sprat |
| 1963 | 50 | 6 | 30 | 21 | 100 | 100 | |
| 1964 | 47 | 11 | 43 | 28 | 29 | 21 | |
| 1965 | 67 | 8 | 30 | 22 | 28 | 11 | |
| 1966 | 60 | 31 | 38 | 62 | 26 | 11 | |
| 1967 | 20 | 100 | 100 | 64 | 22 | 18 | |
| 1968 | 18 | 21 | 18 | 69 | 28 | 9 | |
| 1969 | 78 | 3 | 44 | 35 | 33 | 26 | |
| 1970 | 100 | 47 | 54 | 34 | 26 | 7 | |
| 1971 | 17 | 24 | 68 | 36 | 22 | 16 | |
| 1972 | 35 | 5 | 73 | 41 | 57 | 20 | |
| 1973 | 29 | 36 | 48 | 100 | 49 | 19 | 59 |
| 1974 | 50 | 4 | 68 | 34 | 34 | 7 | 81 |
| 1975 | 24 | 6 | 35 | 25 | 27 | 20 | 35 |
| 1976 | 88 | 12 | 42 | 14 | 54 | 23 | 51 |
| 1977 | 37 | 18 | 46 | 29 | 46 | 7 | 100 |
| 1978 | 34 | 24 | 45 | 42 | 34 | 0.2 | 32 |
| Ratio | 6 | 33 | 6 | 7 | 5 | 500 | 3 |
Fairly clearly with slow growing, low mortality stocks, the yield will come from fish from a number of ages and will thus tend to average out some of the variation in recruitment. On the other hand, fast growing, high mortality stocks will have yields which more nearly vary as recruitment. Thus yields will tend to vary least from year to year on low mortality stocks and on stocks with low variation in recruitment. For such stocks of fish, simple approaches to estimating TACs may be adequate. It may also be possible to make predictions for a number of years. The main safeguards required will be checks that mortality or recruitment levels are not systematically changing. For stocks with a high variation in recruitment and with a small number of ages contributing to the catch, more elaborate methods will be required and predictions will only be reliable for short-time periods. In some cases the shortness of this period may preclude estimating a TAC in sufficient time to be useful. For example, northern North Sea sandeel yields sometimes depends heavily on catches of “O” group fish (in 1979, 61 percent of the total yield) which could only be estimated a few weeks before fishing started. It would therefore be very difficult for scientists to meet to agree on a TAC and administrators to meet to allocate it in this time frame. In general the time lag between data being collected and TACs being set is one year. For example, data are collected in 1982 to be assessed (scientific advice prepared) in 1983 to set the 1984 TAC. Where this is the case, the prospects are not good for setting reasonable TACs on fish stocks with high mortalities and variable recruitments. For such stocks either mortality rates have to be first reduced (if they are man generated) or other methods of management found. Figure 1 shows a schematic picture of this problem. The simplicity or difficulty or impossibility of setting TACs which are precise and accurate is seen as a function of recruitment variability and of the number of ages of fish present in the fishery.
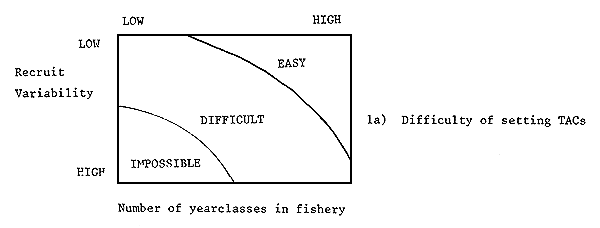
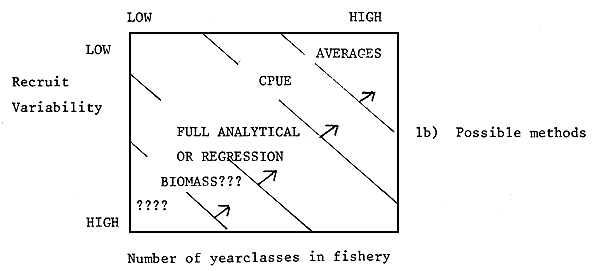
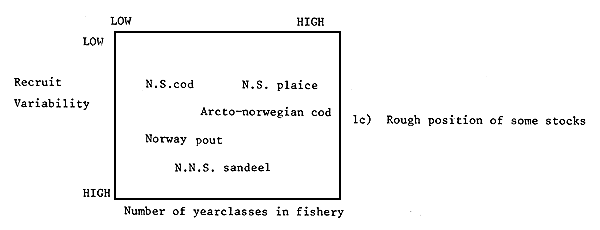
Figure 1
The difficulty of setting TACs, the methods applicable and some typical fish stocks
related to the variability of recruitment and the number of
important year-classes in the fishery
3.2 Estimation method for TACs
Simple approaches :
The simplest approach to TAC estimation is to take a straightforward average of the total catch in the past n years, say over the past five years. Clearly this approach will only be satisfactory if fishing mortality has been stable and if the stock is little affected by recruitment variation, either because variation in recruitment is low or it is averaged over a number of years. While this method is obviously crude, it does reflect the immediate past behaviour of the system. It is possible that some weighted mean of the past five years' data might be a rather better approach since this could give more weight to the most recent years' result. For example, weights of 1/15, 2/15, 3/15, 4/15 and 5/15 might be used to weigh the yields in years t-5, t-4, t-3, t-2, t-1, respectively. Whichever of these methods were used, it would be wise to improve the data available on fishing intensity and recruitment, so that methods which do not rely on the stationarity of recruitment and fishing mortality could be adopted at a later date.
Methods based on catch-per-unit of effort data and effort data :
Where effort and catch-per-unit of effort data are available, predictions of the catch can be made assuming the catch-per-unit of effort in the next year will remain similar to that in the current year. Thus if it is intended to change the fishing mortality (e.g. to Fmax) then the new yield can be predicted as the old level of catch-per-unit of effort times the new effort. This approach was used by ICNAF to set TACs for some species assessed by using production curves (e.g. red fish - McKone, 1978). The method might be a little more elaborated using the approximation given by Walter (1975). This relates catch-per-unit-effort in year i to the fishing effort in year i-1 and thus might be used to improve the estimation of catch-per-unit-effort used to predict the TAC.
Either method could be used with commercial catch-per-unit-effort data or with relative biomass estimates from an annually repeated ground fish survey or an annually repeated acoustic survey. This use of acoustic surveys would not necessarily require an accurate estimate of fish target strength in order for it to be used.
If annual ground-fish survey results are available then it might be possible to use the length distribution of fish caught in the survey to split the catch rate into two or more components, for example, an exploited phase component and a recruiting component. These might be used to form a regression of yield on the two catch rates. A somewhat similar procedure was adopted by the ICES Industrial Fish working group who predicted the 1982 catch of Norway pout from a regression of yield on estimates of one and two-year old fish caught in the international young fish survey which is conducted each year in the North Sea. A somewhat similar approach was proposed for the Arcto-Norwegian cod. Such a method of course supposes that fishing intensity has been stable in the past. In principal mortality rates or estimates of fishing effort might be included in such methods to allow for their changes.
Methods based upon the use of absolute estimates of biomass:
Absolute estimates of biomass could be used to estimate TACs. Obvious sources of such data are acoustic surveys and egg surveys. Results of acoustic surveys of fish stocks can sometimes be made with reasonable accuracy and precision. These might be used directly to estimate the exploitable biomass of the fish stocks and the TAC could then be set at some appropriate proportion of this. One problem with this approach is that the exploitable rather than the total biomass of fish is required. In other words the estimate would have to exclude the biomass of small pre-recruit fish. For species which shoal by size, however, this might be quite possible.
A similar use can be made of egg surveys. First, by using the estimate of total egg production and the fecundity per gram of mature females, to estimate the biomass of mature females and then, using the ratio of catch weight of mature females to all the landings, to gross up the mature female biomass to the total exploitable biomass. Thus again the TAC could be set to give some appropriate proportion of this biomass.
A problem with such TACs would be that they would be estimated on the current biomass and hence could only be used as forward projections if the biomass was unlikely to change, due to low recruitment variability or low mortality rates.
Full Analytical Methods :
In areas such as the North Sea, TACs are set by attempting to estimate the actual numbers at age in the most recent year possible. This is achieved by using VPA or cohort analysis or separable VPA to interpret the catch numbers at age data from past years. This gives estimates of fishing mortality at age and population number at age. These are used to calibrate past estimates of fishing-effort data as fishing mortality and past estimates of recruitment given by research vessel surveys as numbers at age. Current estimates of these factors can then be calibrated to give the current state of the stock including the number of pre-recruit fish. This information can then be used as the basis of a forecast which gives the TAC. This method will be treated in more detail in a paper by Hoydal and is explained in Layman's terms in Pope (1982).
In recent years considerable attention has been given to means of tuning the VPA (e.g. calibrating fishing effort data to give mortality and hence population in the last year). Such methods as the Gamma method, partial exploited biomass method, Saville's method, the Hoydal-Jones method, the Rho (son of Gamma) method (Anon, 1981; Saville, 1981; Laurec and Shepherd, 1982; Armstrong, 1982) are all attempts to make better use of effort data in this context. A preliminary comparison of these methods by an internal working group at the Lowestoft Laboratory (Variations and Fugues for the well-tempered cohort analysis) suggests that most of these methods are reasonable and that discrepancies between them may result more from the effort data used in specific circumstances than the method of calibration adopted. An important scientific problem can therefore be identified which is the need for a method for filtering out bad effort series.
The advantage of the full analytical model is that it attempts to account for all sources of variation in yield. Its disadvantage is that its complex structure might be vulnerable to errors in reporting total catches or discards. It is also vulnerable to poor estimates of catch-at-age data, recruitment or effort data. A potential disadvantage is that it uses data sets which may prove expensive to collect and not be possible for all species. For example, reliable effort data may prove difficult to acquire for shoaling pelagic fish, while age data may not be possible in the tropics. Another problem is that the analytical approach needs a number of years of data before it can be used (perhaps five or more years).
Some recent developments :
Pope (1983) suggests some simplifications to the analytical method for estimating TACs where the proximate goal is for fishing mortality to remain unchanged from the most recent year's level (such TACs may be called status quo TACs). The first of these methods called the leapfrog method can be used with only two years' data and might be of utility when data collecting has only recently been introduced.
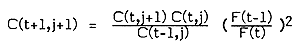
and, 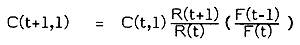
to give catch at age in the TAC year such that F(t+1) = F(t-1). Since fishing mortality estimates appear as ratios, they may be replaced by fishing effort estimates providing these are directly proportional.
The virtue of the method is that it could be used while data for the full analytical method were being accumulated. The main problem is that the equations are likely to be very vulnerable to errors in fishing-effort data because of the use of ratios of effort.
The second of Pope's (1983) method uses the same data set as the full analytical model (i.e. catch at age data for a number of years, fishing effort data and recruitment data). The basis of the method is to interpret the matrix of log-catch ratios - i.e. in (C(i+1, j+1)/C(i,j) - as a two-way analysis of variance (ANOVA). Pope and Shepherd (1982) show this to be an appropriate structure for this matrix. Year effects are then correlated with a formulation of effort data and this is used to extend the log catch ratio matrix by two years (maintaining the year effects and overall mean unchanged). These extended years' log-catch ratios are then used to relate catch at age in the last year for which data are available to the catch at age in the TAC year. An advantage of this method is that no knowledge is required of the level of natural mortality or fishing mortality (indeed these seem somewhat redundant parameters for the purpose of setting a status quo TAC). The method is perhaps most important in showing that setting a status quo TAC is essentially the problem of estimating transfer relationships between catch at age in the last year for which data are available and catch in the TAC year.
Another interesting recent method is the survivors method of Doubleday (1981). This uses what is in effect an exact form of the approximate cohort formulae of Pope (1972). This links population number at the beginning of the year with survivors at the end of the year and catch numbers during the year. This recurrence relationship is: Population = CINT + SINT, where CINT and SINT are an integral of catch and an integral of survivors, both adjusted for accumulated natural mortality. This relationship is then used to generate estimates of population numbers in past years and these are used to calibrate population estimates made from groundfish surveys, using a maximum likelihood procedure. Calibrated population estimates could then of course be projected to give TACs as with the full analytical model. The method could be useful in not using fishing effort data.
Two other methods for calibrating groundfish survey data are shown in Collie and Sissenwine (1982). One method is for when groundfish survey catch at age data and commercial catch at age data are available and the other is for when groundfish survey catch at age data are available but only yield data are available from the commercial catch. Both methods assume a constant catchability coefficient for the research vessel survey data. The method where only total yield data are available from the commercial catch would seem particularly useful because this is a situation where other analytical methods could not be used.
Nielsen (1982) shows a maximum likelihood estimation of fishing mortality based on the separability of mortality into year and age effects. In principle this seems - as did the methods of Doubleday (1976) and Pope (1977) - capable of estimating fishing mortality in the last year using only commercial catch at age data. In practice, however, this is estimated with low precision and thus rather confirms the result of Pope and Shepherd (1982) that there can be little information in the data about mortality in the last year.
New methods that need to be developed :
Analytical methods of TAC estimation still contain a number of scientific problems. Some methods which urgently need development are:
An equivalent to Pope's ANOVA method based on a two or more fleet separable model.
A simpler version of the analytical model based on length data rather than age data. This would be desirable for developing countries where the full analytical model may require more data (particularly age data) than is generally available and for minor or difficult stocks in developed regions.
4. ERRORS IN SOME METHODS OF ESTIMATING TACs
An obvious scientific problem concerned with estimating TACs is how accurate and how precise are the various methods. With regard to accuracy the simple methods make the assumption that the behaviours of fishing mortality and recruitment are simply random and have no trend with time. If either do have a trend with time, the methods will cease to be accurate. For example, if recruitment is collapsing then five-year running means of past catch data will obviously give TACs which are too high. Other methods are obviously susceptible to biases in the data sets used. For example, under reporting of catches, learning processes in fishing effort and trends in fishing power will all potentially upset analytical methods.
The problem of the precision of these various methods is quite difficult. It may be approached in several ways. For example, past predictions may in some cases be compared with the real results as in Macer et al. (1979). It is possible with some methods to see how well they would have worked in past years. For North Sea cod, Table 2 shows the predictions that might have resulted using a five-year running means and contrasting these with the catches that would have occurred had fishing mortality indeed been stabilized at the level of the previous year (adjusted yield). The results suggest a coefficient of variation of about 31 percent. In other words, in some years the results may be out by more than 50 percent. The weighted mean method explained in Section 3 gave much the same result with a coefficient of variation of 28 percent and, in fact, none of these results would seem to improve on setting the TAC equal to the long-term average of the catch.
Table 2
Some possible predictors of catch for North Sea cod and their errors
| Year | Adjusted yields | 5-year running means | Errors | 5-year Weighted means | Errors | Y(t-1) | Errors |
| 68 | 180 | 174 | 6 | 198 | -18 | 245 | -65 |
| 69 | 168 | 207 | -39 | 233 | -65 | 279 | -111 |
| 70 | 318 | 223 | 95 | 299 | 19 | 196 | 122 |
| 71 | 318 | 234 | 84 | 232 | 86 | 230 | 88 |
| 72 | 256 | 256 | 0 | 263 | -7 | 328 | -72 |
| 73 | 299 | 277 | 22 | 295 | 4 | 350 | -51 |
| 74 | 193 | 268 | -75 | 281 | -88 | 236 | -43 |
| 75 | 189 | 269 | -80 | 260 | -71 | 203 | -14 |
| 76 | 198 | 262 | -64 | 234 | -36 | 191 | 7 |
| 77 | 209 | 238 | -29 | 217 | -8 | 212 | -3 |
| 78 | 267 | 208 | -59 | 204 | 63 | 198 | 69 |
| 79 | 247 | 219 | 28 | 231 | 16 | 289 | -42 |
| 80 | 430 | 238 | 192 | 257 | 173 | 299 | 131 |
| Average | 252 | 6.2 | 5.2 | 1.23 | |||
| 75 | 79 | 71 | 78 | ||||
| Coefficient of Variation | 30% | 31% | 28% | 31% |
If the objective of management was to maintain the status quo, then a catch-per-unit-effort adjustment method would simply use the last year's catch to predict this year's catch - using the Walter (1975) approximation would cause some changes - since CPUE and effort would both be the same in the last year and in the predicted year. The results of using Y(t-1) to predict the adjusted catch are shown in Table 2. The coefficient of variation again is about 31 percent.
When acoustic estimates of biomass are used, the precision of the estimated TAC will be equivalent to the estimated precision of the acoustic biomass. Typically the accuracy of acoustic results are quoted in decibels. Often about ± 2 dB's, i.e. about ± 50 percent. Some results, however, have been reported better than this. Jakobsen (1982) shows results that appear to have a coefficient of variation of about 10 percent. To achieve this seems to require an isolated stock heavily concentrated in a known area such as the Icelandic summer spawning herring.
The precision of the 1977 egg survey carried out on the mackerel stock of the south-west of the British Isles gave results which probably had 95 percent confidence regions between +30 percent and - 20 percent of the estimate of egg numbers (Lockwood et al., 1981). The estimate for exploitable biomass would presumably be rather less precise than this, since it would contain additional errors from the fecundity estimate and the mature female proportion in the estimate.
Regression methods of course lend themselves to the estimation of precision. The Norway pout estimates had coefficients of variation of between 15 percent and 25 percent. Using a similar approach for North Sea cod, the results could have a coefficient of variations of about 20 percent, using the international young fish survey results. Not surprisingly a better fit is obtained with the actual numbers at age given by VPA, which could give a result with a coefficient of variation of 10 percent. Whether this discrepancy is due to errors in the total yield estimates and hence catch at age data, or due to changes in the young fish survey, is not clear. In principle a regression approach could be a valuable step between simple methods and the full analytical model.
The precision of the full analytical model is a much more complex question than that of the other methods. Unfortunately, answering this question fully is not easy because the standard practice is both recursive and somewhat subjective. It usually entails the following steps:
Establish data base and update each year
Use VPA or equivalent to interpret past years' catch at age data, subject to assumptions about terminal fishing mortality
Make minor changes to terminal values to obtain consistent interpretation
Calibrate fishing effort data
Use calibrated effort to estimate terminal fishing mortality
Repeat steps (b) and (e) until terminal values are unchanged
Calibrate recruitment indices
Use results from steps above to forecast TACs usually two years ahead
Clearly this calculation route is complex and it is difficult to see how to investigate the variance of the resulting TAC. Several attempts have, however, been made. The problem can be eased by considering only the variability generated in step (h) above. This approach was used by Pope and Garrod (1975) and in a sensitivity analysis shown by Doubleday (1979).
Another alternative is simply to consider how well past TACs have been set (e.g. Macer et al., 1979; Lassen, 1980). Yet another is to set independent working groups to assess the same data as in Garrod, Pope and Knights (1977). This latter approach, of course, only shows the different interpretations that could be put on the same data rather than the true variability. As a short cut, a Monte Carlo simulation approach has much to commend it. Pope and Gray (1983) have adopted such an approach and their results for North Sea plaice, North Sea cod and North Sea sprat are shown in Tables 3 to 11. For each of these three stocks, a simulacrum was constructed which corresponded as closely as possible to the working groups' interpretation, but which had fishing mortality which was separable into year and age effects. This was used to generate exact data sets which were then subjected to different levels of pertubation by random numbers to simulate the inprecise data met with in the real world. A series of TACs were then generated at each error level for each data set to give a 4 × 4 × 4 factorial design. TACs were calculated to meet three possible proximate goals. These were, where F(t) is the fishing mortality in the last year for which data are available and year t+2 the TAC year:
F(t+2) = F(t)
F(t+2) = Fmax
F(t+2) = 0.5
In all cases F(t+1) = F(t).
In practice additional important sources of variation arise from weight at age (Rivard, 1981) and uncertainty about fishing mortality in year T+1. What is particularly interesting about the results is that variability seems to be far less when F(t+2) = F(t) than the results for other proximate goals. This is a result of some internal compensations in the method when this proximate goal is chosen. It is also least subject to effort generated errors, which is fortunate since this is less capable of change than the other error sources. Thus, fairly clearly, the precision of TACs is a function of the amount of sampling input to the stock and also of the choice of proximate goal. There are this problems here for both scientists and administrators to solve.
Table 3
Coefficients of variation of 1982 North Sea plaice TAC
resulting from various coefficients of variation of inputs
to TACs, where F(1982) = F(1980)
| Fishery effort data | Catch at age data | Recruitment data | |||
| (%) | (%) | 0% | 15% | 30% | 60% |
| 0 | 0 | 0 | 3 | 6 | 12 |
| 10 | 5 | 6 | 8 | 13 | |
| 20 | 10 | 11 | 12 | 16 | |
| 40 | 22 | 23 | 24 | 26 | |
| 15 | 0 | 8 | 9 | 11 | 15 |
| 10 | 10 | 11 | 12* | 16 | |
| 20 | 14 | 14 | 16 | 19 | |
| 40 | 26 | 26 | 27 | 29 | |
| 30 | 0 | 17 | 17 | 18 | 21 |
| 10 | 18 | 18 | 19 | 22 | |
| 20 | 21 | 22 | 22 | 25 | |
| 40 | 32 | 33 | 34 | 37 | |
| 60 | 0 | 29 | 29 | 30 | 32 |
| 10 | 29 | 29 | 30 | 32 | |
| 20 | 31 | 31 | 31 | 34 | |
| 40 | 37 | 37 | 38 | 39 | |
* Highlighted cell is authors' personal estimate of current situation
Table 4
Coefficients of variation of 1982 North Sea plaice TAC
resulting from various coefficients of variation of inputs
to TACs, where F(1982) = Fmax
| Fishery effort data | Catch at age data | Recruitment data | |||
| (%) | (%) | 0% | 15% | 30% | 60% |
| 0 | 0 | 0 | 3 | 6 | 12 |
| 10 | 8 | 9 | 10 | 15 | |
| 20 | 14 | 15 | 16 | 19 | |
| 40 | 30 | 31 | 31 | 34 | |
| 15 | 0 | 23 | 23 | 24 | 26 |
| 10 | 25 | 26 | 27* | 29 | |
| 20 | 30 | 30 | 31 | 33 | |
| 40 | 45 | 46 | 47 | 49 | |
| 30 | 0 | 48 | 49 | 49 | 51 |
| 10 | 52 | 53 | 53 | 56 | |
| 20 | 57 | 58 | 58 | 61 | |
| 40 | 80 | 80 | 81 | 84 | |
| 60 | 0 | 112 | 112 | 113 | 115 |
| 10 | 115 | 115 | 116 | 118 | |
| 20 | 122 | 122 | 123 | 124 | |
| 40 | 136 | 136 | 137 | 138 | |
* Highlighted cell is authors' personal estimate of current situation
Table 5
Coefficients of variation of 1982 North Sea plaice TAC
resulting from various coefficients of variation of inputs
to TACs, where F(1982) = 0.5
| Fishery effort data | Catch at age data | Recruitment data | |||
| (%) | (%) | 0% | 15% | 30% | 60% |
| 0 | 0 | 0 | 3 | 6 | 12 |
| 10 | 7 | 7 | 9 | 14 | |
| 20 | 13 | 14 | 15 | 18 | |
| 40 | 28 | 29 | 30 | 32 | |
| 15 | 0 | 23 | 23 | 24 | 27 |
| 10 | 24 | 25 | 25* | 28 | |
| 20 | 28 | 29 | 29 | 32 | |
| 40 | 43 | 44 | 44 | 47 | |
| 30 | 0 | 48 | 48 | 49 | 51 |
| 10 | 50 | 50 | 51 | 53 | |
| 20 | 56 | 56 | 57 | 59 | |
| 40 | 77 | 77 | 78 | 80 | |
| 60 | 0 | 112 | 112 | 113 | 114 |
| 10 | 113 | 113 | 114 | 115 | |
| 20 | 119 | 119 | 120 | 122 | |
| 40 | 132 | 132 | 133 | 135 | |
* Highlighted cell is authors' personal estimate of current situation
Table 6
Coefficients of variation of 1982 North Sea cod TAC
resulting from various coefficients of variation of inputs
to TACs, where F(1982) = F(1980)
| Fishery effort data | Catch at age data | Recruitment data | |||
| (%) | (%) | 0% | 15% | 30% | 60% |
| 0 | 0 | 0 | 12 | 23 | 43 |
| 10 | 2 | 12 | 23 | 44 | |
| 20 | 5 | 13 | 24 | 44 | |
| 40 | 10 | 15 | 25 | 45 | |
| 15 | 0 | 13 | 17 | 26 | 45 |
| 10 | 13 | 18 | 27* | 45 | |
| 20 | 14 | 18 | 27 | 46 | |
| 40 | 17 | 21 | 29 | 47 | |
| 30 | 0 | 23 | 26 | 32 | 48 |
| 10 | 24 | 26 | 33 | 48 | |
| 20 | 24 | 27 | 33 | 49 | |
| 40 | 27 | 30 | 35 | 50 | |
| 60 | 0 | 34 | 36 | 37 | 49 |
| 10 | 35 | 36 | 39 | 49 | |
| 20 | 36 | 37 | 40 | 50 | |
| 40 | 40 | 39 | 44 | 53 | |
* Highlighted cell is authors' personal estimate of current situation
Table 7
Coefficients of variation of 1982 North Sea cod TAC
resulting from various coefficients of variation of inputs
to TACs, where F(1982) = Fmax
| Fishery effort data | Catch at age data | Recruitment data | |||
| (%) | (%) | 0% | 15% | 30% | 60% |
| 0 | 0 | 0 | 12 | 24 | 44 |
| 10 | 5 | 13 | 24 | 44 | |
| 20 | 9 | 15 | 25 | 45 | |
| 40 | 13 | 18 | 27 | 47 | |
| 15 | 0 | 22 | 25 | 33 | 51 |
| 10 | 22 | 26 | 33* | 51 | |
| 20 | 24 | 27 | 34 | 52 | |
| 40 | 27 | 29 | 36 | 54 | |
| 30 | 0 | 43 | 45 | 50 | 65 |
| 10 | 43 | 45 | 50 | 65 | |
| 20 | 44 | 46 | 51 | 66 | |
| 40 | 48 | 50 | 55 | 69 | |
| 60 | 0 | 84 | 85 | 87 | 96 |
| 10 | 85 | 85 | 88 | 97 | |
| 20 | 87 | 87 | 90 | 99 | |
| 40 | 95 | 96 | 98 | 107 | |
* Highlighted cell is authors' personal estimate of current situation
Table 8
Coefficients of variation of 1982 North Sea cod TAC
resulting from various coefficients of variation of inputs
to TACs, where F(1982) = 0.5
| Fishery effort data | Catch at age data | Recruitment data | |||
| (%) | (%) | 0% | 15% | 30% | 60% |
| 0 | 0 | 0 | 12 | 23 | 44 |
| 10 | 4 | 13 | 24 | 44 | |
| 20 | 9 | 15 | 25 | 45 | |
| 40 | 17 | 21 | 29 | 47 | |
| 15 | 0 | 22 | 25 | 33 | 51 |
| 10 | 22 | 26 | 33* | 51 | |
| 20 | 24 | 27 | 34 | 51 | |
| 40 | 29 | 31 | 37 | 54 | |
| 30 | 0 | 43 | 45 | 50 | 65 |
| 10 | 43 | 45 | 50 | 65 | |
| 20 | 44 | 46 | 51 | 66 | |
| 40 | 49 | 51 | 55 | 69 | |
| 60 | 0 | 84 | 85 | 88 | 96 |
| 10 | 85 | 86 | 88 | 97 | |
| 20 | 87 | 88 | 90 | 99 | |
| 40 | 95 | 96 | 99 | 107 | |
* Highlighted cell is authors' personal estimate of current situation
Table 9
Coefficients of variation of 1982 North Sea sprat TAC
resulting from various coefficients of variation of inputs
to TACs, where F(1982) = F(1980)
| Fishery effort data | Catch at age data | Recruitment data | |||
| (%) | (%) | 0% | 15% | 30% | 60% |
| 0 | 0 | 0 | 9 | 19 | 40 |
| 10 | 2 | 9 | 19 | 40 | |
| 20 | 4 | 10 | 19 | 40 | |
| 40 | 9 | 12 | 20 | 41 | |
| 15 | 0 | 1 | 9 | 19 | 40 |
| 10 | 3 | 9 | 19 | 40 | |
| 20 | 5 | 10 | 19 | 40 | |
| 40 | 9 | 12 | 20 | 40 | |
| 30 | 0 | 5 | 10 | 19 | 39 |
| 10 | 5 | 10 | 19 | 39 | |
| 20 | 7 | 11 | 19 | 39* | |
| 40 | 11 | 14 | 21 | 40 | |
| 60 | 0 | 15 | 18 | 24 | 40 |
| 10 | 16 | 18 | 24 | 40 | |
| 20 | 17 | 19 | 25 | 40 | |
| 40 | 20 | 22 | 27 | 42 | |
* Highlighted cell is authors' personal estimate of current situation
Table 10
Coefficients of variation of 1982 North Sea sprat TAC
resulting from various coefficients of variation of inputs
to TACs, where F(1982) = Fmax
| Fishery effort data | Catch at age data | Recruitment data | |||
| (%) | (%) | 0% | 15% | 30% | 60% |
| 0 | 0 | 0 | 9 | 20 | 42 |
| 10 | 3 | 10 | 20 | 42 | |
| 20 | 6 | 11 | 20 | 42 | |
| 40 | 12 | 15 | 22 | 43 | |
| 15 | 0 | 11 | 14 | 22 | 43 |
| 10 | 11 | 14 | 22 | 43 | |
| 20 | 12 | 15 | 23 | 43 | |
| 40 | 16 | 18 | 25 | 44 | |
| 30 | 0 | 26 | 28 | 33 | 50 |
| 10 | 26 | 28 | 33 | 50 | |
| 20 | 27 | 29 | 34 | 50* | |
| 40 | 29 | 31 | 35 | 51 | |
| 60 | 0 | 87 | 88 | 91 | 100 |
| 10 | 87 | 88 | 91 | 100 | |
| 20 | 88 | 89 | 92 | 101 | |
| 40 | 90 | 92 | 95 | 103 | |
* Highlighted cell is authors' personal estimate of current situation
Table 11
Coefficients of variation of 1982 North Sea sprat TAC
resulting from various coefficients of variation of inputs
to TACs, where F(1982) = 0.5
| Fishery effort data | Catch at age data | Recruitment data | |||
| (%) | (%) | 0% | 15% | 30% | 60% |
| 0 | 0 | 0 | 9 | 18 | 39 |
| 10 | 3 | 9 | 18 | 39 | |
| 20 | 7 | 11 | 19 | 39 | |
| 40 | 14 | 16 | 22 | 40 | |
| 15 | 0 | 13 | 15 | 22 | 34 |
| 10 | 13 | 16 | 23 | 41 | |
| 20 | 14 | 17 | 23 | 42 | |
| 40 | 19 | 21 | 26 | 40 | |
| 30 | 0 | 32 | 34 | 38 | 51 |
| 10 | 33 | 34 | 38 | 52 | |
| 20 | 33 | 35 | 39 | 53* | |
| 40 | 36 | 37 | 41 | 54 | |
| 60 | 0 | 103 | 104 | 107 | 115 |
| 10 | 103 | 104 | 107 | 115 | |
| 20 | 104 | 105 | 108 | 115 | |
| 40 | 107 | 108 | 110 | 118 | |
* Highlighted cell is authors' personal estimate of current situation
5. SCIENTIFIC PROBLEMS CAUSED BY TECHNOLOGICAL INTERACTIONS AND MULTI-SPECIES FISHERIES
Many fisheries use methods of capture which can and do catch several different species at the same time. The tactics of fishermen may also very reasonably include diversion from fishing the main species sought during a fishing trip to fishing a secondary species when an attractive concentration of this species is found. For example, Lowestoft fishermen usually fish for plaice with one set of gear but may transfer to another gear if a good concentration of cod is encountered and market conditions are right. Thus, whether by accidental capture or deliberate diversion, it is normal for a fishery which generates fishing mortality on one species to generate by-catch fishing mortalities on other species.
These by-catch mortalities can frequently cause conflicts of interest between fisheries for different species. This is particularly likely to happen when the by-catch is generated from intensive small-mesh fisheries for small fast-growing species. By-catches in these fisheries may well be juveniles of other species which should be left to grow on.
Some examples of such conflicts are haddock, which was caught as a by-catch in the silver hake fishery on Georges Bank, groundfish caught as by-catch in Norway-pout fisheries in the North Sea, herring caught as by-catch in sprat fisheries and groundfish caught in inshore shrimp fisheries in the South China Sea area and elsewhere. In shrimp fisheries, discard rates for groundfish as high as 95 percent have been recorded.
Such problems may sometimes be alleviated by restricting the areas in which small-mesh gears may be used or possibly by gear modifications aimed at releasing some species while capturing the species at which fishing is directed.
Where these palliatives are unsuccessful in resolving conflicts, then there may be a need for mortalities generated on both fisheries to be adjusted. There may, for example, be a need for the fishery for the fat-growing species to be reduced to a somewhat lower level than would be optimal were only that fishery considered. Similarly, the species caught as a by-catch may need to have a higher than optimal level of mortality. Thus, even if no biological interactions occur between species, such technological interactions may cause a change of objectives for both species. The choice of new objectives of course falls on the managers, since there must be a compromise between the yield to be had from two different fisheries, and therefore based on economic rather than yield considerations. The scientific problem is, of course, elucidating the consequences.
The problem can become particularly acute where managers wish to prevent any fishing on a species in order to allow it to recover from severe depletion. Strictly enforced such regulations might be very restrictive on other viable fisheries. The haddock on Georges Bank and the North Sea herring are both examples of this problem.
Where by-catch fishing mortality is causing the virtual extinction of a species, the problem can be particularly difficult to solve. An example of the problem is given by skates and rays which Holden (1977) shows can be fished down at quite low fishing mortality rates due to their low fecundity. The common skate (Raja batis) is particularly at risk and Brander (1981) shows that this species has effectively disappeared from the Irish Sea. It may be that, for this species to be conserved, fishing mortality rates on all Irish Sea demersal fish might need to be reduced.
Certainly this example does point out how difficult the choice of objectives for a mixed fishery may be.
The problems posed to scientists by mixed fisheries are often concerned with the extent that by-catch rates might be moderated by given regulations and the consequences of change in mortality rates. A common problem is the definition of main species sought and the true level of by-catch. Unless fairly disaggregated species catch data are recorded for each fleet, the by-catch rate can be apparently increased by the results of separate fisheries being merged.
One solution to the mixed fishery problem tried on Georges Bank was the imposition of a second-tier TAC on all species combined. This was to restrict catches to prevent mortalities on some species being generated to the detriment of other species. In practice, this was difficult for scientists to calculate with any certainty, and the problem might perhaps be better solved by political and economic compromises between the conflicting fisheries.
Although the problems of mixed fisheries show the need for setting objectives which compromise between species, the need is even more obvious where species interactions occur. For example, predation by fish of one species on juveniles of another species is likely to produce a distinct change in the yield of the second species. Clearly its yield may be affected by fishing mortality levels on the first species. An example of this is the increase in the abundance of squid in the Gulf of Thailand which appears to have resulted from a general decrease in the abundance of most other fish species, due to fishing pressure. The scientific problem might be seen as being to define the consequences of particular levels of fishing on each species. This would, however, at our present state of knowledge, be wildly ambitious. The immediate problem may therefore be to indicate possible effects of directions of change in mortality for various key species. A good review of the scientific problems of multi-species fisheries may be found in Mercer (1982) which contains a series of papers given at a Workshop on Multi-species Fisheries held at St. Johns, Newfoundland in 1980.
At a more immediate level, multi-species predation effects do not appear to cause very much of a problem for setting TACs to maintain current mortality levels. This is because much of the predation would seem to be on ‘O’ group fish (at least in the North Sea) and thus research vessel recruitment surveys and commercial catch-at-age data are not much affected by varying levels of predation mortality (Pope and Knights, 1982). Table 12 shows mortalities calculated by their method for a number of North Sea fish stocks.
Table 12 Natural mortalities generated by two runs of the modified cohort analysis
| Species | Run | A g e s | ||||||||||||||
| 0 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | ||
| Cod | 1 | 0.975 | 0.206 | 0.104 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | ||||||
| 2 | 0.838 | 0.227 | 0.11 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | |||||||
| Haddock | 1 | 0.847 | 0.403 | 0.242 | 0.115 | 0.108 | 0.104 | 0.102 | 0.1 | 0.1 | ||||||
| 2 | 0.776 | 0.407 | 0.261 | 0.133 | 0.120 | 0.111 | 0.104 | 0.1 | 0.1 | |||||||
| Whiting | 1 | 1.114 | 0.405 | 0.149 | 0.131 | 0.131 | 0.131 | 0.118 | 0.109 | |||||||
| 2 | 0.976 | 0.419 | 0.191 | 0.167 | 0.167 | 0.167 | 0.141 | 0.121 | ||||||||
| Saithe | 1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | |||
| 2 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | ||||
| Herring | 1 | 0.407 | 0.271 | 0.260 | 0.220 | 0.218 | 0.219 | 0.193 | 0.143 | 0.143 | ||||||
| 2 | 0.248 | 0.291 | 0.279 | 0.261 | 0.259 | 0.259 | 0.234 | 0.142 | 0.142 | |||||||
| Plaice F | 1 | 0.1 | 0.117 | 0.109 | 0.103 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 |
| 2 | 0.1 | 0.121 | 0.118 | 0.108 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | |
| Plaice M | 1 | 0.1 | 0.117 | 0.109 | 0.106 | 0.103 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | |||||
| 2 | 0.1 | 0.121 | 0.118 | 0.108 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | |
6. REFERENCES
Armstrong, D.W. and R.M. Cook, 1982 A proposal for the use of catch per unit effort data for the assessment of terminal F in V.P.A. ICES CM. 1982/G:53 (mimeo)
Brander, K., 1981 Disapperance of common skate, Raia batis, from the Irish Sea. Nature,Lond., (5801):48–9
Collie, J.S. and M.P. Sissenwine, 1982 Estimating population size from relative abundance data measured with error. ICES CM. 1982/G:34 (mimeo)
Doubleday, W.G., 1976 A least squares approach to analysing catch at age data. Res.Bull.ICNAF, (12): 69–81
Doubleday, W.G., 1979 A sensitivity analysis of selected catch projections. ICES CM. 1979/G:53:7 p. (mimeo)
Doubleday, W.G., 1981 A method of estimating the abundance of survivors of an exploited fish population using commercial catch at age and research vessel abundance indices. Can.Spec.Publ. Fish.Aquat.Sci., (58):164–78
Garrod, D.J., J.G. Pope and B.J. Knights, 1977 The ICES stock evaluation course: an experiment in replicate assessments. ICES CM. 1977/F:20:12 p. (mimeo)
Holden, M.J., 1977 Elasmobranchs. In Fish population dynamics, edited by J.A. Gulland. London, Wiley, Interscience, 187–215 pp.
ICES, 1981 Report of the ad hoc Working Group on the Use of effort data in assessments. ICES CM. 1981/G:5 (mimeo)
Jakobssen, J., 1983 Echo surveying of the Icelandic summer spawning herring, 1973–1982. FAO Fish.Rep., (300)
Lassen, H., 1980 On the quality of the TACs recommended through ICES. ICES SM. 1980/G:39 (mimeo)
Laurec, A. and J.G. Shepherd, 1982 On the analysis of catch and effort data. ICES CM. 1982/G:8 (mimeo)
Lockwood, S.J., J.H. Nichols and W.A. Dawson, 1981 The estimation of a mackerel (Scomber scombrus) spawning stock size by plankton survey. J.Plankton Res., 3(2):217–33
McKone, W.D., 1978 Assessment of divisions 3LN redfish. ICNAF Res.Doc. 78/VI/46:10 p.
Macer, C.T., B.W. Jones and R.C.A. Bannister, 1979 Factors affecting the accuracy of catch predictions in some demersal fish stocks. ICES CM. 1979/G:34:5 p. (mimeo)
Mercer, M.C. (ed.), 1982 Multispecies approaches to fisheries management advice. Can.Spec.Publ.Fish. Aquat.Sci., (59):169 p.
Nielsen, N.A., 1982 Estimation of VPA-Parameters in a model of restricted fishing mortalities. ICES CM. 1982/G:50 (mimeo)
Pope, J.G., 1972 An investigation of the accuracy of virtual population analysis using cohort analysis. Res.Bull.ICNAF, (9):65–74
Pope, J.G., 1977 Estimation of fishing mortality, its precision and implications for the management of fisheries. In Fisheries mathematics, edited by J.H. Steele. London, Academic Press, pp. 63–76
Pope, J.G., 1979 Stock assessment in multispecies fisheries, with special reference to the trawl fishery in the Gulf of Thailand. Manila, South China Sea Fisheries Development Programme, SCS/DEV/79/19:106 p.
Pope, J.G., 1981 Practical guidelines for the precision of assessment data. ICES CM. 1981/G:13:11 p. (mimeo)
Pope, J.G., 1982 Background to scientific advice on fisheries management. Lab.Leafl.Dir. Fish.Res.G.B., (54):26 p.
Pope, J.G., 1983 Analogies to the status quo TACs: their nature and variance. Can.Spec.Publ.Fish. Aquat.Sci. (in press)
Pope, J.G. and D.J. Garrod, 1975 Sources of error in catch and effort quota regulations with particular reference to variations in the catchability coefficient. Res.Bull.ICNAF, (11):17–30
Pope, J.G. and D.F. Gray, 1983 Practical guidelines for the precision of assessment data. Can.Spec.Publ. Fish.Aquat.Sci. (in press)
Pope, J.G. and B.J. Knights, 1982 Simple models of predation in multi-species for considering the estimation of fishing mortality and its effects. Can.Spec.Publ.Aquat.Sci., (59):64-9 pp.
Pope, J.G. and J.G. Shepherd, 1982 A simple method for the consistent interpretation of catch-at-age data. J.Cons.CIEM, 40:176–84
Rivard, D., 1981 Catch projections and their relation to sampling error of research surveys. Can.Spec. Publ.Fish.Aquat.Sci., (58):93–109
Saville, A., 1981 Comments by ACFM on the report of the North Sea Roundfish Working Group. ICES CM. 1980/G:80:8 p. (mimeo)
Shepherd, J.G., 1982 Two measures of overall fishing mortality. ICES CM. 1982/28:11 p. (mimeo)
Waller, G.G., 1975 Graphical methods for estimating parameters in simple models of fisheries. ICNAF Res.Doc. 51 Ser.No.3530 (mimeo)
![]()
![]()
![]()